光合成の理論計算
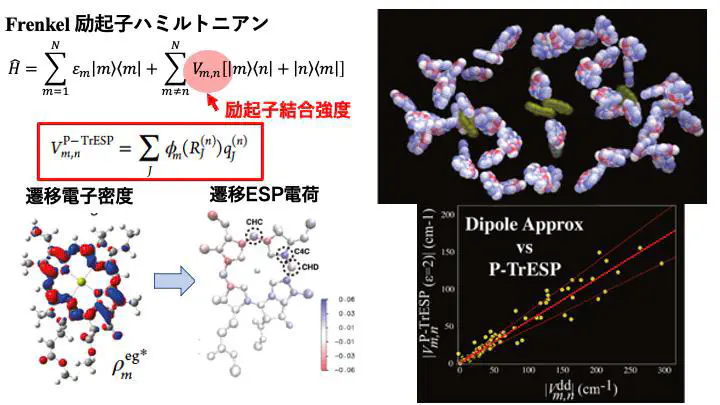
光合成の光捕集機構を理論的に解析するには、光合成反応中心や光捕集アンテナ蛋白質の構造情報から、 アンテナ色素のサイトエネルギーと励起子相互作用を計算し、励起子ハミルトニアンを構築する必要があります。
我々はこれまで、非酸素発生I型光合成反応中心(RC)として、2017年に初めてX線結晶構造が報告された ヘリオバクテリアRC(hRC)に注目した研究を行ってきました。 酸素発生を行う光合成生物は、それぞれ光化学系I(PSI)、光化学系II(PSII)と呼ばれる I型とII型の両方の光合成反応中心を持っており、 X線結晶構造についても既に報告されています。
- シアノバクテリアのPSIがヘテロダイマーなのに対し、hRCはホモダイマー構造を持つこと
- hRCは周辺光捕集蛋白質を持たず、PSIのコア蛋白質にChlorophyll aが96個存在するのに対し、 hRC中では54 Bacteriochlorophyll g (BChl g)、4 BChl g’と、アンテナ色素数も少ないこと
などから、hRC特有の光捕集機構が働いていると考えれます。
そこで我々は、hRCとPSIの光捕集機能の差異を調べるため、色素間の励起子結合強度を、 量子化学計算に基づく遷移ESP電荷(TrESP)法を用いて計算しました。 その結果、hRCとPSIでは、似た箇所に配置されている色素でも、 それらの励起子結合強度は全く異なる性質を持つことを見出しました。 その原因を探るため、hRCとPSIの持つ色素を計算機上で仮想的に置換する実験を行い、 hRCが持つ特殊なクロロフィル(BChl g)の電子状態とその空間的配置の2つの要因が、 結合強度分布の違いを生み出すことを明らかにしました。
現在、アカリオクロリス生物や緑色硫黄光合成細菌の反応中心に対してもこれらの手法を適用することで、 I型光合成反応中心の光捕集機構や電子伝達機構を統一的に理解する研究を進めています。
(2020).
Excitonic Coupling on a Heliobacterial Symmetrical Type-I Reaction Center: Comparison with Photosystem I.
The Journal of Physical Chemistry B Vol. 124, num.2, pp. 389–403 (2020).
(2021).
Comparison between the Light-Harvesting Mechanisms of Type-I Photosynthetic Reaction Centers of Heliobacteria and Photosystem I: Pigment Site Energy Distribution and Exciton State.
The Journal of Physical Chemistry B Vol. 125, num.15, pp. 3727-3738 (2021).